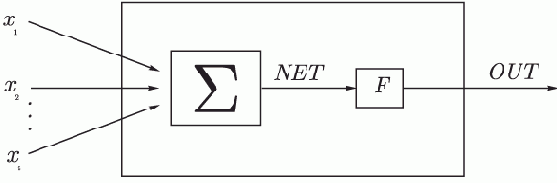
Искусственный нейрон имитирует в первом приближении свойства биологического нейрона. На вход искусственного нейрона поступает некоторое множество сигналов, каждый из которых является выходом другого нейрона. Каждый вход умножается на соответствующий вес, аналогичный синаптической силе, и все произведения суммируются, определяя уровень активации нейрона.
Рис. 1.2.
На рис. 1.2 представлена модель, реализующая эту идею. Множество входных сигналов, обозначенных
, поступает на искусственный нейрон. Эти входные сигналы, в совокупности обозначаемые вектором
, соответствуют сигналам, приходящим в синапсы биологического нейрона. Каждый сигнал умножается на соответствующий вес
, и поступает на суммирующий блок, обозначенный
. Каждый вес соответствует "силе" одной биологической синаптической связи. (Множество весов в совокупности обозначается вектором
.) Суммирующий блок, соответствующий телу биологического элемента, складывает взвешенные входы алгебраически, создавая выход, который мы будем называть
. В векторных обозначениях это может быть компактно записано следующим образом:
Сигнал
далее, как правило, преобразуется активационной функцией
и дает выходной нейронный сигнал
. Активационная функция может быть обычной линейной функцией
где
— константа, пороговой функцией
где
— некоторая постоянная пороговая величина, или же функцией, более точно моделирующей нелинейную передаточную характеристику биологического нейрона и предоставляющей нейронной сети большие возможности.
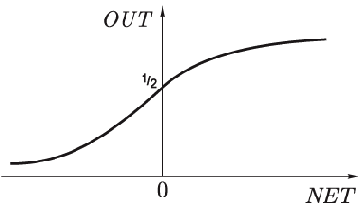
Рис. 1.3.
На рис. 1.2 блок, обозначенный
, принимает сигнал
и выдает сигнал
. Если блок
сужает диапазон изменения величины
так, что при любых значениях
значения
принадлежат некоторому конечному интервалу, то
называется "сжимающей" функцией. В качестве "сжимающей" функции часто используется логистическая или "сигмоидальная" (S-образная) функция, показанная на рис. 1.3. Эта функция математически выражается как
. Таким образом,
По аналогии с электронными системами активационную функцию можно считать нелинейной усилительной характеристикой искусственного нейрона.
Коэффициент усиления вычисляется как отношение приращения величины
к вызвавшему его небольшому приращению величины
. Он выражается наклоном кривой при определенном уровне возбуждения и изменяется от малых значений при больших отрицательных возбуждениях (кривая почти горизонтальна) до максимального значения при нулевом возбуждении и снова уменьшается, когда возбуждение становится большим положительным. С. Гроссберг (1973) обнаружил, что подобная нелинейная характеристика решает поставленную им дилемму шумового насыщения. Каким образом одна и та же сеть может обрабатывать как слабые, так и сильные сигналы? Слабые сигналы нуждаются в большом сетевом усилении, чтобы дать пригодный к использованию выходной сигнал. Однако усилительные каскады с большими коэффициентами усиления могут привести к насыщению выхода шумами усилителей (случайными флуктуациями), которые присутствуют в любой физически реализованной сети. Сильные входные сигналы, в свою очередь, также будут приводить к насыщению усилительных каскадов, исключая возможность полезного использования выхода. Центральная область логистической функции, имеющая большой коэффициент усиления, решает проблему обработки слабых сигналов, в то время как области с падающим усилением на положительном и отрицательном концах подходят для больших возбуждений. Таким образом, нейрон функционирует с большим усилением в широком диапазоне уровня входного сигнала
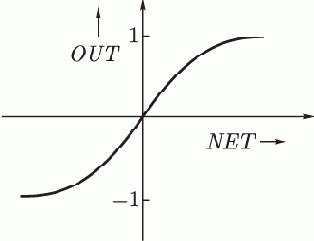
Другой широко используемой активационной функцией является гиперболический тангенс. По форме она сходна с логистической функцией и часто используется биологами в качестве математической модели активации нервной клетки. В качестве активационной функции искусственной нейронной сети она записывается следующим образом:
Рис. 1.4.
Подобно логистической функции гиперболический тангенс является S-образной функцией, но он симметричен относительно начала координат, и в точке
значение выходного сигнала
равно нулю (см. рис. 1.4). В отличие от логистической функции, гиперболический тангенс принимает значения различных знаков, и это его свойство применяется для целого ряда сетей.
Рассмотренная простая модель искусственного нейрона игнорирует многие свойства своего биологического двойника. Например, она не принимает во внимание задержки во времени, которые воздействуют на динамику системы. Входные сигналы сразу же порождают выходной сигнал. И, что более важно, она не учитывает воздействий функции частотной модуляции или синхронизирующей функции биологического нейрона, которые ряд исследователей считают решающими в нервной деятельности естественного мозга.
Несмотря на эти ограничения, сети, построенные из таких нейронов, обнаруживают свойства, сильно напоминающие биологическую систему. Только время и исследования смогут ответить на вопрос, являются ли подобные совпадения случайными или же они есть следствие того, что в модели верно схвачены важнейшие черты биологического нейрона.